“First do no harm”
“Primum non nocere”
The long-term clinical effectiveness of Lysis is unparalleled compared to other therapies. High patient satisfaction and your reputation go hand in hand with outcomes.

Lab
Learn from luminary physicians at a Racz Lab. Gain hands-on experience, learn the nuances, and safety protocols of Pain Medicine techniques. Engage and network with fellow physicians. Get vital reimbursement lectures and practice management insights. Join us for unparalleled education in interventional pain medicine.
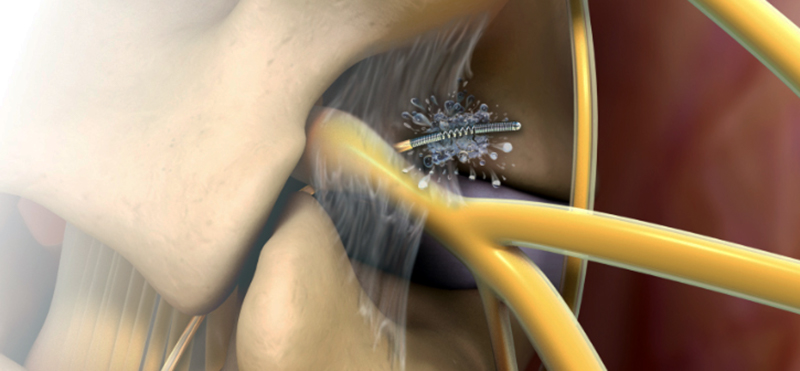
Library
Unlock your full potential in Interventional Pain Medicine at the Racz Library. Discover comprehensive education and training by Dr. Gabor B. Racz. Enhance your skills with lectures, videos, case studies, and interactive modules. Learn from luminary physicians and gain expertise in cervical and thoracic pain management. Elevate your practice with comprehensive online education.
Training
Take your training to a higher level and connect with a Racz Certified Trainer. Browse our international database to find the perfect match and receive 1-on-1 training in person. Unlock your full potential with expert guidance and take your practice to the next level.
Coming Soon!

We are proud of our mission
At RaczEdu, our mission is to revolutionize the field of Interventional Pain Medicine by imparting the knowledge and skills necessary to provide effective pain management solutions. We strive to create a global community of well-trained physicians who are passionate about delivering compassionate care to patients suffering from chronic pain. Through our comprehensive educational resources and training programs, we aim to elevate the standard of pain management worldwide.











